
Czynniki wpływające na procesy generatywne u zbóż ozimych.
Rośliny w ciągu swojego życia przechodzą przez kolejne fazy rozwojowe od siewki do dojrzałości pełnej, gdy roślina wytwarza nasiona. Fazy rozwojowe to nic innego jak tworzenie nowych organów np.: liści, korzeni, łodyg, węzłów (nazywamy je organami wegetatywnymi), oraz elementów kwiatostanów takie jak pręciki, pylniki, słupek, płatki korony, czy plewki u zbóż (nazywamy je organami generatywnymi – biorą udział w tworzeniu nasion). Za tworzenie każdego z tych elementów odpowiada inny zestaw genów, które muszą być aktywowane w odpowiedniej kolejności i odpowiednim miejscu rośliny.
Co sprawia, że geny „wiedzą”, kiedy mają zacząć pracować, a kiedy mają być uśpione?
Sprawcą jest kod DNA, który działa jak program informatyczny, dla każdego gatunku odmiennie. Program jest „odpalany” (inaczej mówiąc uruchamiany) przez czynniki środowiskowe. Tymi czynnikami są temperatura, światło, dostępność do wody. Jednak nie sama obecność wymienionych czynników uruchamia geny odpowiedzialne za tworzenie poszczególnych organów rośliny lecz zmiany w odpowiednim zakresie. Na przykład: spadek czy wzrost temperatury poniżej określonej wartości, zmiana długości dnia do poziomu indukcyjnego, zmiana widma światła słonecznego zarówno w poszczególnych porach roku jak i podczas doby.
Dlaczego kod DNA zawarty w genach działa jak program informatyczny ? bo geny są uruchamiane warunkowo. Na przykład roślina przejdzie z fazy tworzenia liści do fazy tworzenia kwiatostanu jeśli = pod warunkiem, że wystąpi okres chłodu który zablokuje geny represorowe kwitnienia, tj. takie geny, które blokują geny odpowiedzialne za tworzenie kwiatostanu. To jest jeden z wielu przykładów pokazujących jak roślina zarządza swoim rozwojem.
Rośliny zbożowe w toku milionów lat ewolucji dostosowywały się do warunków klimatycznych określonej szerokości geograficznej.
W każdej szerokości geograficznej inny czynnik najsilniej wpływa na rozwój rośliny. Na przykład w warunkach równikowych i podzwrotnikowych, gdzie nie ma dobowych i sezonowych zmian długości dnia i temperatury, kluczowa jest dostępność wody. W klimacie gorącym dominują gatunki, które kwitną po osiągnięciu określonej fazy rozwoju, np. kukurydza. Jeśli brakuje wody roślina nie wytwarza kolejnych liści, a więc nie wchodzi w następne fazy rozwoju.
W klimacie umiarkowanym mrozy występujące zimą pokrywają się z krótkim dniem. W takich warunkach zadomowiły się tylko takie gatunki i odmiany, które są wstanie dostosować czas kwitnienia do wiosennej pory roku. Ważne dla zbóż ozimych i ciekawe jest to, że z dwóch wymienionych czynników klimatycznych długość dnia zmienia się co roku tak samo, bo zależy od położenia planety Ziemia w układzie słonecznym. Natomiast przebieg temperatury jest każdego roku nieco inny.
Gatunki zbóż ozimych wysiane na przełomie lata i jesieni nie dadzą się zwieść pojawiającej się co kilka lat wysokiej temperaturze na początku jesieni. Nie tworzą kwiatostanów nawet gdy osiągną wystarczającą do tego fazę rozwoju. Dzieje się tak dlatego, że są wstanie rejestrować, (co roku tak samo) zmniejszającą się długości dnia jesienią. Ona jednoznacznie wskazuje na zbliżającą się zimę. W ten sposób wieloczynnikowa zależność rozwoju generatywnego pozwala roślinie integrować informacje płynące ze zmian warunków atmosferycznych i astronomicznych.
Zboża ozime zakwitną dopiero w 2 połowie maja. Aby nie wymarzły podczas zimy muszą aktywować geny blokujące niektóre z naturalnych procesów: najpierw indukcji, potem inicjacji i w końcu różnicowania i rozwoju kwiatostanu tj. kłosa.
Procesy prowadzące do kwitnienia zbóż ozimych są kilku etapowe i bardzo złożone. Niemal co roku pojawiają się w prasie fachowej doniesienia o odkryciu kolejnych genów biorących udział w tym procesie. W tym artykule jedynie zasygnalizujemy najlepiej poznane procesy.
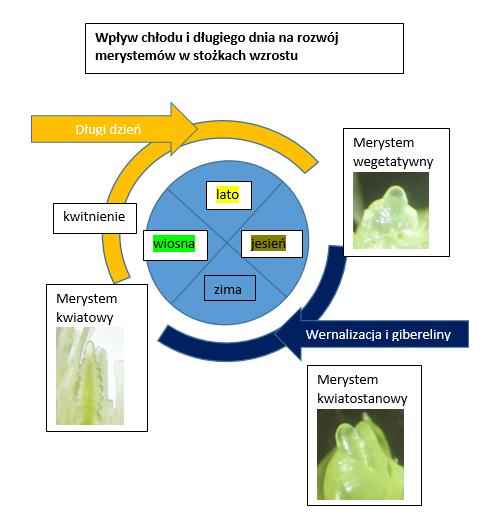
Indukcja kwitnienia polega na aktywacji ( chłód i długi dzień) genów znoszących blokadę wysłania sygnałów z liści do merystemów. Sygnałami są białka, które po dotarciu do celu ( merystemu) przyczyniają się do zmian morfologicznych, które prowadzą do powstania zaczątku kwiatostanu . Ten proces przemiany merystemu z wegetatywnego w generatywny nazywany jest inicjacją kwitnienia. Procesy przemiany merystemu prowadzą po wielu etapach do powstania plewek, pylników, słupków kwiatowych. Gdyby nie indukcja i inicjacja, z merystemów powstały by liście.
Jesień
Wiadomo od dawna, że jesienią niska temperatura w zakresie 10-0*C indukuje procesy generatywne( rozpoczyna proces tworzenia kwiatostanu) przez zniesienie blokady, którą stanowi gen VRN2. Proces ten nazywany jest wernalizacją, bądź jaryzacją. Skutki zniesienia blokady będą widoczne dopiero wiosną, gdy dzień zacznie się wydłużać powyżej 12 godz. Wówczas uruchomi się uwolniony od blokady genu VRN2 szlak indukcji kwitnienia zależny od czasu naświetlenia liści (indukcyjnego fotoperiodu).
Najkrócej ujmując dzieje się tak dlatego, że chłód zmienia konformację czyli budowę przestrzenną DNA i białek histonowych, a tym samym uaktywnia gen VRN1. Jego produktem jest białko mające zdolność blokowania genu VRN2, który wcześniej blokował szlak indukcji kwitnienia zależny od fotoperiodu. Rysunek poniżej.
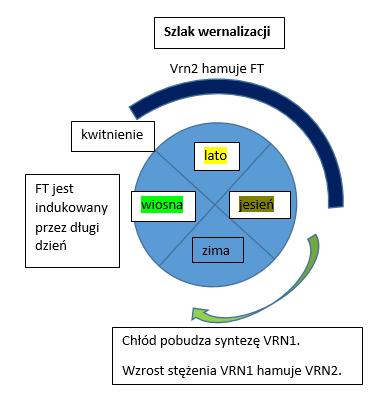
Minimalny czas naświetlania liści, aby spowodować indukcję – (zapoczątkowanie morfogenezy kwiatowej) wynosi u roślin zbożowych 12 godz. Dla szerokości geograficznej Polski indukcyjny długi dzień kończy się w końcu września a zaczyna ok. 20 marca. U zbóż ozimych do końca września jest aktywny gen blokujący szlak indukcji kwitnienia zależny od długiego dnia. W przeciwnym razie u wcześnie wysianych zbóż, indukcyjny długi dzień rozpoczął by proces generatywny zbyt wcześnie.
W październiku dzień jest zbyt krótki by światło w ciągu dnia mogło indukować kwitnienie, a mimo to już na początku grudnia można zaobserwować na dobrze wykształconych rozkrzewieniach zaczątki kłosów będące w fazie wałeczkowania. Świadczy to niezbicie o przejściu procesu indukcji ale także trwającej inicjacji kwitnienia (zmiany morfologiczne w stożku wzrostu). Przyczyną jest drugi efekt działania produktu białkowego genu VRN1. Jego białko aktywuje geny w komórkach stożków wzrostu odpowiedzialne za indukcję kwitnienia, najpierw SOC1, AGL24, SVP…… , a te wiosną aktywują kolejne tzw. geny tożsamości kwiatowej: LFY, grupę genów APETALA odpowiedzialnych za rozwój kwiatów w kwiatostanie.
Proces indukcji kwitnienia jest wspomagany jesienią przez gibereliny.
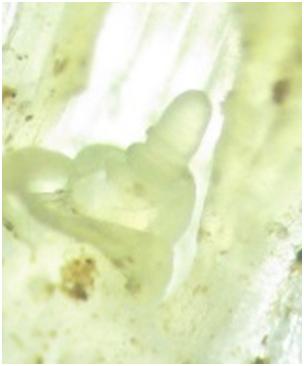
Dwie grupy bioaktywnych giberelin o odmiennym sposobie działania spływają z liści do mereystemów. Jedne z nich GA1 i GA4 silnie wpływają na wydłużanie komórek. Druga grupa GA5 i GA6 nie wydłuża komórek natomiast aktywuje geny inicjujące zmiany morfologiczne w stożkach wzrostu widoczne na fot. 1 i 2.

Fot. 1 Merystem przed inicjacją. Fot: Mariusz Anioła

Fot. 2 Merystem kwiatostanowy widoczne początkowe zmiany morfologiczne. Fot: Mariusz Anioła
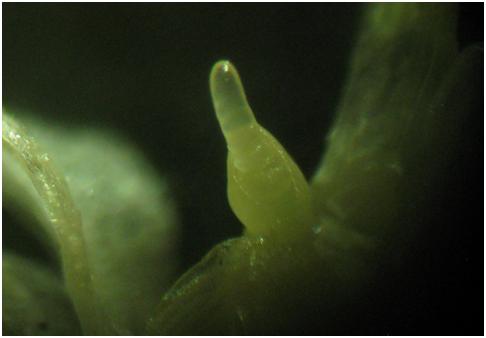
Fot. 3 Inicjacja stożka wzrostu. Fot: Mariusz Anioła

Fot.4 Inicjacja stożka wzrostu. Fot: Mariusz Anioła
Fot. 3 i 4 Warunki indukcyjne tj. chłód jesienią zaindukowały – uaktywniły geny VRN 1(szlak wernalizacyjny), oraz gen odpowiedzialny za biosyntezę enzymu 2 oxydaza giberelinowa (szlak giberelinowy), które aktywowały geny LFY, SOC 1 odpowiedzialne za inicjację kwiatostanu w stożku wzrostu.
Wałeczki na wydłużonym, wąskim merystemie są dowodem na trwający proces inicjacji kwiatostanu fot 5.
Fot. 5 Wyraźnie widoczne wałeczki z których powstaną kłoski – pięterka. Fot: Mariusz Anioła
Udział giberelin w jesiennej indukcji kwitnienia.
Gdy nastaną niskie temperatury następuje demetylacja białek histonowych i w konsekwencji aktywacja genu odpowiedzialnego za produkcję enzymu GA2ox w strefie podwierzchołkowej merystemu. Ten enzym nie jest wstanie rozłożyć, ważnych dla indukcji kwitnienia GA5 i GA6 natomiast rozkłada pozostałe gibereliny np.: GA1, GA4. Przewaga ilościowa giberelin indukujących kwitnienie nie trwa długo, ale raz zapoczątkowany proces indukcji jest już nieodwracalny.
Wiosną gdy temperatura wzrośnie i długość dnia zrówna się z nocą tj. ok. 20 marca GA1 i GA4 będzie docierała do merystemu w większych ilościach. Wówczas te hormony odegrają ważną rolę w różnicowaniu kwiatów.
Niska temperatura nie tylko jest pierwotnym czynnikiem który uruchamia indukcję kwitnienia jesienią ale także uruchamia procesy przygotowujące roślinę do spoczynku zimowego. Proces indukcji kwitnienia jesienią i spoczynku są ze sobą powiązane.
Czynniki zewnętrzne niesprzyjające wzrostowi jednocześnie sprzyjają indukcji i inicjacji kwitnienia. Czynniki zewnętrzne sprzyjające wzrostowi sprzyjają już zainicjowanemu wcześniej kwiatostanowi do jego rozwoju.
Wiosna

Fot. 6 Wiosenna indukcja kwitnienia. Fot: Mariusz Anioła
Wiosenna indukcja kwitnienia zależna od fotoperiodu jest możliwa dzięki zablokowaniu genu VRN2 podczas jesiennej vernalizacji. Ten gen blokował inny ważny gen, którego produkt białkowy FT1 przedostaje się z liści do stożków wzrostu i tam aktywuje geny odpowiedzialne za przemianę merystemu wegetatywnego w generatywny. (rys 1)
Wiosną po 20 marca ponad 12 godzinny dzień sprzyja wzrostowi stężenia barwników (fitochromy i kryptochromy) aktywujących geny (GI, CO). Te z kolei aktywują gen FT1, będący przekaźnikiem między czynnikami zewnętrznymi, a merystemem w stożkach wzrostu.
Barwniki aktywujące gen FT1 aktywują również gen biosyntezy enzymu 20 oxydazy giberelinowej. Wraz z innymi enzymami 3 oxydaza giberelinowa i 2 oxydaza giberelinowa powodują wzrost stężenia bioaktywnych giberelin. Tak jak jesienią tak i wiosną GA5 i GA6 indukują merystemy powstające podczas wiosennego krzewienia. Wiosną GA1 i GA4 wpływają na wydłużanie międzywęźli oraz wspomagają morfogenezę (rozwój) kwiatów (fot. 7, 8, 9, 10)
-
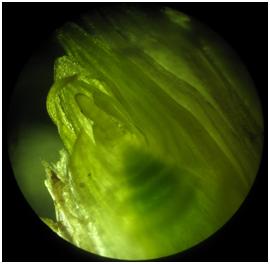
- Fot. 7 Data 01.03.2017
-

- Fot. 8. Data 10.03.2017
-
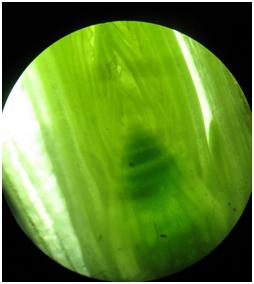
- Fot. 9 Data 19.03.2017
-
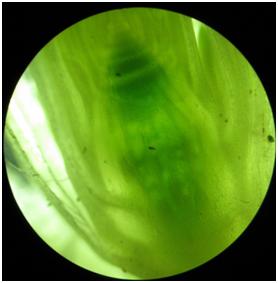
- Fot. 10 Data 19.03.2017
Fot: Mariusz Anioła
Boczne rozgałęzienia (górna strzałka) na zaczątku kłosa to merystemy kwiatowe. Są pobudzane do wzrostu przez gibereliny GA1 i GA4 – Fot. 11

Fot. 11 Merystemy kwiatowe. Fot: Mariusz Anioła
05.04.2017 główne rozgałęzienie. Widoczne wydłużanie się międzywęźli to skutek działania giberelin powstających w liściach. Międzywęźla powyżej są jeszcze bardzo krótkie, to dlatego, że liście które z nich wyrastają są jeszcze skryte przed światłem wewnątrz pochew starszych liści i nie powstaje w nich odpowiednia ilość giberelin – fot. 12

Fot. 12 Główne rozgałęzienia. Fot: Mariusz Anioła
Wydłużanie się międzywęźli odbywa się dzięki wydłużaniu poszczególnych komórek pod wpływem giberelin głównie GA1 i GA4, oraz dzięki pomnażaniu ich w węzłach. Węzły są miejscem gdzie znajdują się komórki merystematyczne mające zdolność do podziałów. Intensywność podziałów (zwiększania liczby komórek) jest zależna od ilości dopływających cytokinin ze stożków wzrostu korzeni, oraz proporcji do pozostałych hormonów głównie giberelin a także auksyn, etylenu i wielu innych.
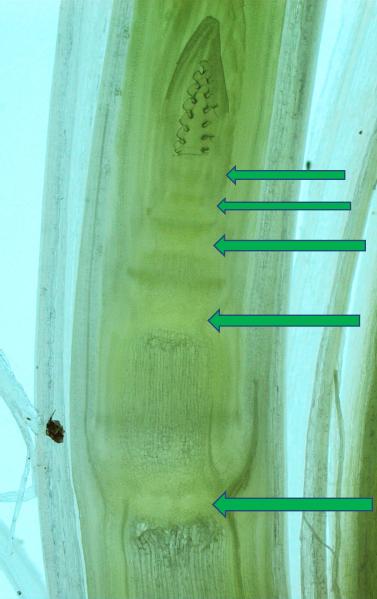
Fot.13 Strzałki pokazują węzły z komórkami merystematycznymi. W merystemach następują mitotyczne podziały komórkowe stymulowane przez hormony głównie cytokininy. Fot: Mariusz Anioła

Fot. 14 Wydłużanie się międzywęźli. Data 14.04.2017. Fot: Mariusz Anioła

Fot.15 Kolejny etap różnicowania się kłosa i wydłużania międzywęźli. Fot: Mariusz Anioła
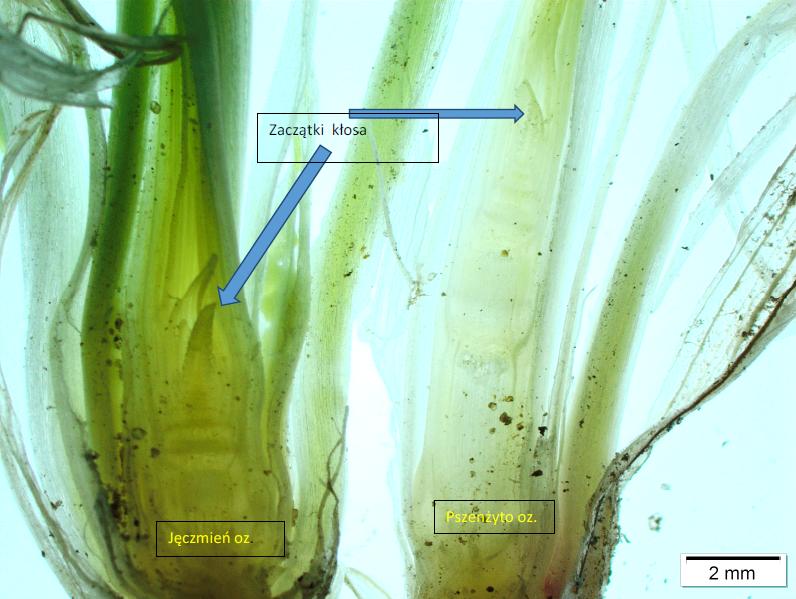
Fot. 16 Jęczmień i pszenżyto wysiano w tym samym terminie. Wydłużanie międzywęźli jest mniej intensywne u jęczmienia niż u pszenżyta. Data 24.04.2017 r. Fot: Mariusz Anioła

Fot.17 Gibereliny GA1 i GA4 grają dużą rolę w rozwoju kwiatów w kłosie. Data 14.04.2017, Fot: Mariusz Anioła
Praktyczne uwagi:
Niska temperatura jesienią uruchamia proces wernalizacji i wraz z giberelinami indukuje a potem inicjuje jeszcze przed zimą zaczątki kłosów w rozkrzewieniach. Dla praktyki ważny jest optymalny termin siewu taki aby roślina mogła wytworzyć 3 główne, silne rozkrzewienia jeszcze jesienią. Wówczas jeszcze przed zimą chłód aktywuje geny kwitnienia w stożkach wzrostu. Im dłużej temperatura jest w granicach 0-10*C tym więcej nowo powstających w merystemach komórek ulega indukcji. Im więcej komórek jest zaindukowanych tym dłuższy kłos.
Wiosną czasu na indukcję i inicjację jest niewiele bo już przed 20 marca zaczyna się nowy etap rozwoju zainicjowanych wcześniej stożków wzrostu. Nie przybywa kolejnych kłosków a jedynie rozwijają się ich merystemy. Jeśli zima skończy się późno to kłosonośnych wiosennych rozkrzewień będzie bardzo mało a kłosy będą krótkie.
Pośród zbóż ozimych pszenica krzewi się najlepiej. Jednak i u tego gatunku są duże różnice wśród odmian. Odmiany dobrze się krzewiące to takie, które są wstanie syntetyzować większe ilości cytokinin. To hormony tworzące się głównie w korzeniach a po dotarciu do stożków wzrostu stymulują komórki merystematyczne do intensywnych podziałów mitotycznych. Im wcześniej rośliny wznowią wegetację tym dłużej cytokininy stymulują krzewienie. Dominacja cytokinin kończy się gdy wzrasta poziom giberelin wydłużających źdźbło. Poziom giberelin rośnie od początku marca, ale nasila się od momentu gdy długość dnia zrównuję się z nocą. (rozchodzenie się węzłów jest dowodem na aktywność giberelin). Wzrost temperatury, długości dnia i pojawianie się kolejnych liści powoduje ciągły wzrost giberelin, których stężenie hamuje działanie cytokinin. Kończy się krzewienie i zaczyna intensywny wzrost na długość – faza strzelania w źdźbło.
Dla słabo rozkrzewionych zbóż ozimych zaleca się stosowanie preparatów zawierających CCC (chlorek chlorocholiny). Ta substancja zatrzymuje syntezę giberelin na wcześniejszym etapie niż inne preparaty stosowane do skracania źdźbła. Tym samym pozwala bez przeszkód działać cytokininom i rozkrzewiać zboża, jeśli CCC zostanie podana w fazie BBCH 25-27.
CCC i kilka innych substancji aktywnych (chlorek chloromekwatu, Etefon i inne) używa się do skracania źdźbła w ściśle określonych fazach dla każdej z substancji. Każda z nich hamuje syntezę giberelin wydłużających międzywęźla.
Stosowanie inhibitorów giberelin do rozkrzewiania i skracania źdźbła narusza równowagę hormonalną w roślinie. W fazie „wielkiego okresu” nie zaleca się stosowania inhibitorów min. dlatego, że wówczas gibereliny są zaangażowane w rozwój męskiej części kwiatu.
Podsumowanie:
Potencjał plonotwórczy zbóż jest kształtowany na bardzo wczesnych etapach rozwoju. Procesy prowadzące do powstawania elementów kłosa (zamiast liści) są wzbudzane (paradoksalnie) przez dość niesprzyjające warunki środowiskowe. Jednak kolejne etapy wymagają już bardziej sprzyjających warunków klimatycznych. Warunki środowiskowe temperatura, długość dnia wpływają na uaktywnienie jednych genów i hamowanie innych. Od składu genów, i ich stopnia dominacji zależy reakcja gatunku czy odmiany na czynniki klimatyczne, np. kwitnienie. Np. odmiany o bardziej recesywnym genie vrn1 mają wyższe wymagania co do okresu wernalizacji niż odmiany posiadające gen vrn1 o większej dominacji. Te ostatnie powinny być siane w optymalnym terminie. Lepsza znajomość procesów mających wpływ na tworzenie plonu pozwoli na uświadomienie sobie, który element agrotechniki mógł wpłynąć na ograniczenie plonu. Powyższy artykuł pokazuje jak bardzo skomplikowany i wieloetapowy jest rozwój roślin.

Jestem w trakcie robienia projektu i ten wpis bardzo mi się przydał. Wielkie dzięki! Pozdrawiam 😉